

Bioenergetics rarely makes the news. But one molecule—2,4-Dinitrophenol—forces us to look directly into the engine room of human metabolism. In the volatile world of DNP mitochondria research, this compound breaks every rule of how we create energy.
Most textbooks treat DNP as a toxin or a historical mistake from the 1930s. That view misses the point. After spending 15 years looking at metabolic research, I see something else. We aren’t just looking at a banned substance; we are looking at the most aggressive form of mitochondrial uncoupling known to science.
The papers from Nature or the NIH give you the chemistry. But they often fail to explain the visceral reality of why this mechanism is so potent and so dangerous.
This is not an endorsement. This is a deep-dive into the mechanics of your cells when the brakes get cut. We’re going to look at the physics of the proton gradient, why researchers are currently trying to “tame” this molecule for diabetes, and the razor-thin margin where biology fails.
What the Textbooks Miss
When you read standard literature on 2,4-Dinitrophenol, three critical details often get buried in the jargon. We need to pull them out.
- The “Runaway Train” Loop: Most papers describe what happens (uncoupling) but miss why it kills. The body has no off-switch for chemically induced uncoupling.
- Targeting the Liver: The new wave of research (like the 2020 Nature Communications study) flips the script from “systemic poison” to “targeted therapy.”
- Bypassing the Turbine: We need to visualize exactly how DNP makes the ATP Synthase enzyme useless, turning mitochondria into simple heaters.
1. The Engine Room: How Mitochondrial Uncoupling Actually Works
To understand DNP, you have to understand the “Chemiosmotic Theory.”
Think of a dam. Your mitochondria pump protons (water) behind a wall (the inner membrane). This builds pressure. To relieve that pressure, the protons have to flow back through a specific turbine: ATP Synthase. As they rush through, the turbine spins, and the cell creates ATP (energy).
The DNP Sabotage
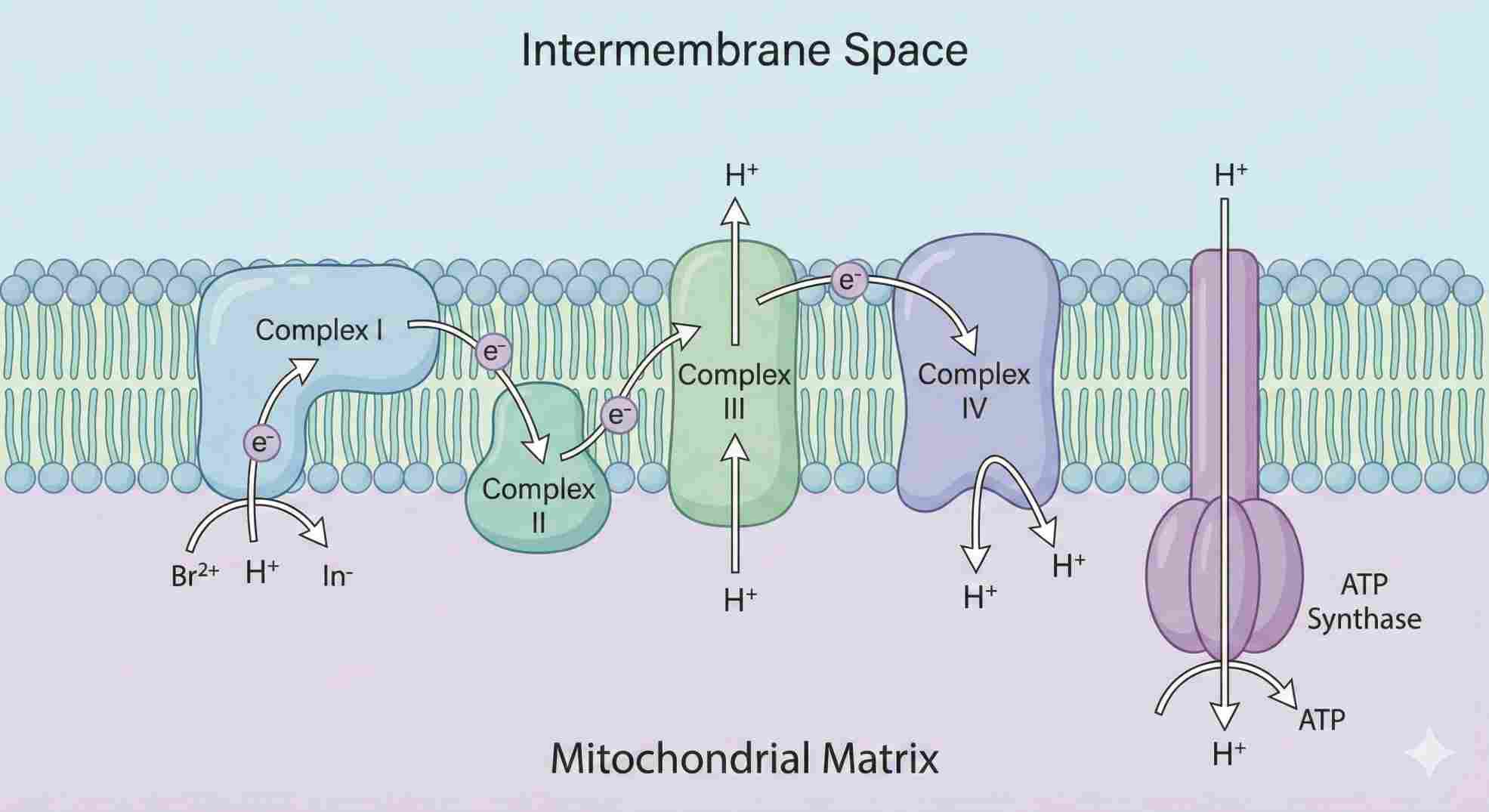
DNP acts like a smuggler. It’s a lipophilic weak acid. It grabs a proton on the high-pressure side, slips right through the membrane wall (ignoring the turbine entirely), and drops the proton on the other side.
The result?
- Pressure Drop: The proton gradient collapses.
- Panic: The cell senses the loss of pressure. It screams for more fuel. It burns glucose and fat furiously to pump those protons back out.
- Waste: Since the protons just leak back in via DNP, the turbine never spins. No ATP is made. The energy from that burned fat doesn’t move a muscle. It releases solely as heat.
Note: This isn’t just “burning calories.” This is metabolic sabotage. Your cells are shoveling coal into a furnace with the windows open.
2. The Heat Death: Why Uncoupling Has No Ceiling
Medical literature usually cites “hyperthermia” as a side effect. That’s putting it mildly.
Normally, if you exercise hard, you get hot. You stop running, and your body cools down. Your mitochondria obey signals to slow down.
DNP ignores signals.
Because the uncoupling is chemical, not hormonal, the mitochondria cannot stop burning fuel. As long as the molecule is there, the leak continues.
- Normal Fever: The body sets the thermostat to 101°F.
- DNP Hyperthermia: The furnace burns until the house catches fire. The body cooks from the inside out. This explains the rigor mortis seen in older case studies—the muscles run out of ATP and lock up instantly at death.
Data: DNP vs. Normal Metabolism
We looked at bioenergetics data to compare standard states against DNP-induced states.
(Scroll right to view full table)
| Metabolic State | ATP Production | Heat Production | Oxygen Consumption | Limiting Factor |
|---|---|---|---|---|
| Resting | Normal | Low | Low | ATP Demand |
| Intense Exercise | High | High | High | Oxygen Delivery |
| DNP Uncoupling | Zero/Low | Extreme | Maximum | Fuel Availability |
3. The Pivot: From Toxin to Medicine?
Here is the most critical gap in the current narrative. If DNP is so deadly, why did Nature Communications (2020) and Diabetes (2019) publish papers on it?
Researchers are now engineering Controlled-Release Mitochondrial Protonophores (CRMP).
The “Hepatic” Strategy
The danger of DNP is that it hits every cell—heart, brain, muscle. But what if you could restrict the uncoupling only to the liver?
Recent studies looked at “liver-targeted” DNP derivatives. These modified molecules aim to:
- Reverse Fatty Liver: By increasing fat oxidation specifically in liver cells.
- Fix Insulin Resistance: By lowering the fat burden on organs, insulin sensitivity improves.
- Avoid Hyperthermia: By keeping the uncoupling local, the systemic temperature spike never happens.
This suggests the mechanism isn’t the villain; the delivery system is.
Research Note: The Diabetes journal study noted that a controlled-release version reversed diabetes in rats without the toxicity of free DNP. This is a massive shift in how we view metabolic treatments.
4. The Margin of Error
In toxicology reports, Dinitrophenol stands out because it lacks a therapeutic window. Most drugs have a wide gap between the dose that works and the dose that kills. DNP doesn’t.
The “Rule of Three” Paradox
In the 1930s, doctors prescribed DNP. They quickly realized a terrifying math problem:
- Dose A: Metabolic rate up by 20% (Weight loss).
- Dose A x 3: Metabolic rate up by 500% (Death).
The difference between a “miracle” and the morgue is a multiplier of three. Few substances on earth carry such a steep curve.
(Scroll right to view full table)
| Dosage Multiplier | Physiological Effect | Metabolic Consequence |
|---|---|---|
| Baseline (Dose A) | Therapeutic Window (Historical) | +20% Metabolic Rate Increase |
| Toxic (Dose A x 3) | Fatal Hyperthermia | +500% Metabolic Rate Increase |
| Comparison: Caffeine | High Safety Margin | Requires ~50-100x dose for lethality |
Respecting the Fire
We started by looking at a molecule often dismissed as a diet pill. We end by looking at the fundamental limits of biology.
DNP proves that the human body is an engine built for efficiency, not waste. When we break that efficiency through mitochondrial uncoupling, we force fat loss, but we risk the very mechanism that keeps us alive.
The future of this field lies not in the raw chemical, but in the precision engineering seen in the latest papers. Until science perfects the “targeted leak,” the mitochondria remain a powerful engine that demands respect.
If you are looking for safe, established ways to optimize this engine rather than risk it, we explore the legal and healthy side of bioenergetics in our main guide: The Mitochondrial Renaissance: A Deep Dive into Mitochondrial Support Supplements.
If you are researching bioenergetics, remember: The proton gradient is the battery of life. Short-circuiting it is the most effective way to drain the battery—and often, the life along with it.

Ready to see real changes in the mirror? Try Mitolyn risk-free for 90 days and lock in today’s special discount before it expires
Frequently Asked Questions:
Does DNP stop the electron transport chain (ETC)?
No. Cyanide stops the chain. DNP actually lets the chain run faster. It disconnects the electron transport from the ATP creation. The electrons flow, oxygen gets used, but no useful energy is stored.
What happens to Basal Metabolic Rate (BMR)?
DNP artificially inflates BMR. Exercise raises metabolic rate via demand; DNP raises it via inefficiency. Historical data showed BMR increases of 30-50% in patients, a rate you can’t really sustain naturally without running a marathon every day.
Can antioxidants stop DNP toxicity?
No. The toxicity is heat, not oxidative stress. While free radicals are produced, the primary killer is hyperthermia and running out of ATP. No vitamin can cool down a cell that is chemically uncoupled.
Is DNP legal for research?
DNP is widely used in in vitro (test tube) research to study mitochondria. It is a standard tool for scientists to measure “maximal respiration” in cells. However, it is strictly banned for human consumption.